
How a Nerve Fires
Or: how a soggy-biocritter thinks in electricity

Before we can talk about how the brain learns to ignore things (which it does with astonishing enthusiasm), we should first understand how a nerve cell actually speaks.
Because here is the first surprise:
Your thoughts are not made of “energy.” They are not “vibrations.” They are not mystical emanations of consciousness.
They are salt.
Well. Salt, water, and exquisitely organised lipid membranes.
Welcome to the electrochemical life of the soggy-biocritter.
A Neuron at Rest Is Not Resting
A nerve cell sitting quietly is not neutral. It is charged. Literally.
Inside the neuron is slightly negative relative to the outside — about minus seventy millivolts. That may not sound like much, but at cellular scale it is dramatic. It is tension. Poise. Readiness.
How does it maintain this tiny battery?
By doing something profoundly unglamorous and energetically expensive: pumping sodium out and potassium in. All day. All night. No applause.
The sodium–potassium pump is the cellular equivalent of someone endlessly rearranging deck chairs on the Titanic to maintain a charge gradient.
Why?
Because life runs on gradients.
A neuron at rest is a cliff edge of possibility.
Threshold: Commit or Don’t
Neurons are not dramatic creatures. They don’t fire at every whisper.
Small inputs arrive. A little sodium leaks in. A tiny depolarisation occurs. Nothing spectacular.
But if enough small nudges accumulate — if the membrane potential crosses a certain threshold — something decisive happens.
Voltage-gated sodium channels snap open.
And then there is no half-measure.
This is what we call an action potential.
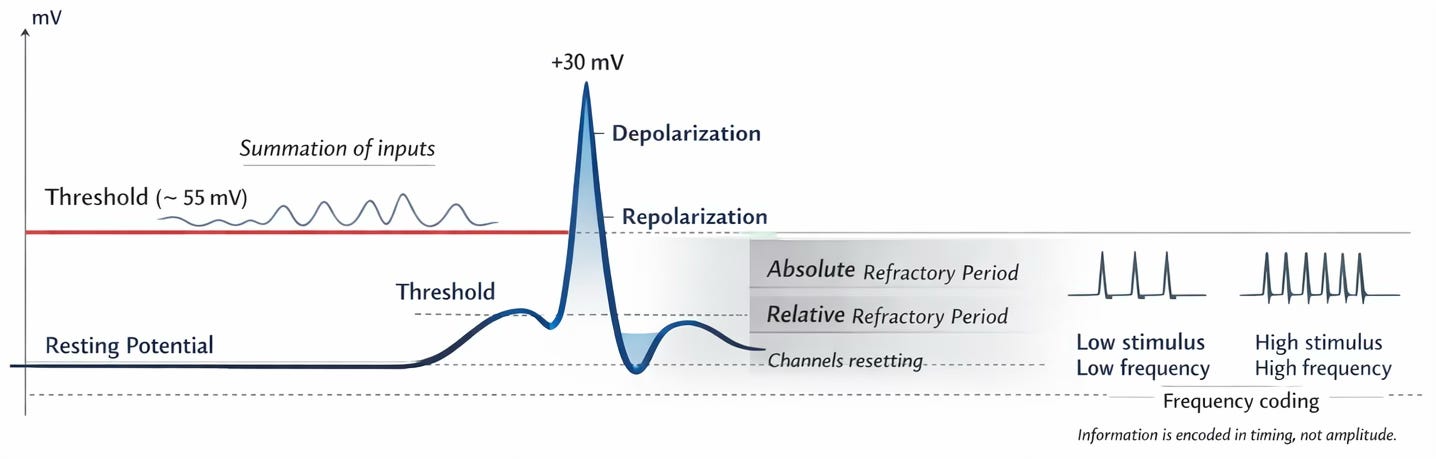
It is all-or-none. No “maybe.” No “partial enlightenment.”
Either the neuron commits — or it does not.
Biology, in this case, is refreshingly binary.
The Action Potential: A Traveling Storm
When threshold is reached, sodium floods in. The inside of the cell becomes rapidly positive.
Then, just as rapidly, potassium channels open and the cell repolarises.
This entire event takes a few milliseconds.
But here is the elegant part:
The action potential does not flow like electricity in a copper wire. It regenerates itself along the membrane. Each segment triggers the next.
It is more like a wave in a stadium crowd than a current in a cable.
No electron is travelling from your toe to your brain.
The membrane itself is performing a carefully choreographed voltage dance.
And it does so thousands of times per second if required.
Your nervous system is not quiet.
It is a very busy salt orchestra.
When Electricity Becomes Chemistry
Eventually the action potential reaches the end of the axon.
Here, something extraordinary happens.
The electrical signal opens calcium channels. Calcium rushes in. Vesicles fuse with the membrane. Neurotransmitter is released into the synapse.
The signal becomes chemical.
One neuron whispers across a microscopic gap to the next.
And here — right here — is where modulation lives.
Because synapses are adjustable.
They can release more transmitter.
Or less.
They can become more sensitive.
Or less.
The volume knob of the nervous system lives at the synapse.
Remember that. We will need it later.
Frequency Is the Real Language
Neurons do not shout louder by producing “bigger” action potentials.
An action potential is always roughly the same size.
Instead, they communicate by frequency.
More important stimulus? Fire more often.
Less important? Fire less often.
This is closer to FM radio than to a light switch.
Which means your perception of the world is not about amplitude alone.
It is about pattern. Timing. Rate.
Your nervous system is a temporal code machine.
Neurons do not care about static levels.
They care about change.
Because they are already firing. They are already idling.
A thermoreceptor is not asleep until warmth arrives. It is ticking along at a baseline rate.
When temperature rises, firing frequency increases.
When temperature falls, firing frequency decreases.
The brain does not measure “warm.”
It measures “warmer than before.”
The nervous system is less like a thermometer and more like a Difference Engine.
Not an absolute reader.
A comparator.
Signal minus baseline.
The Cost of Firing
Every action potential costs energy.
Ions move down gradients. Pumps must restore them. ATP, the molecular currency in soggy-biocritters, is consumed.
The brain, though only a small fraction of body mass, consumes a disproportionate amount of metabolic energy.
Thinking is expensive.
Attention is expensive.
Spikes are expensive.
Which raises a practical biological question:
If something in the environment is harmless and predictable…
should we keep spending glucose on it?
If the nervous system is fundamentally a difference detector — a comparator — then what happens when there is no difference?
When the stimulus stops changing?
At that point, the system has a choice.
Continue to amplify a static signal…
Or recalibrate the baseline itself.
And that recalibration is what we call habituation.
A Final Thought Before We Move On
A neuron does not “decide” in the psychological sense.
It obeys physics and chemistry.
But when billions of neurons interact — modulating thresholds, adjusting synapses, altering firing frequency — something remarkable emerges:
Selective awareness.
The brain is not a camera.
It is an editor built on ion gradients.
And next time, we will explore one of its most elegant editing tools:
Habituation.
That was Signal Over Noise today.